Typologie nationale des biocénoses benthiques de Méditerranée (NatHab-Med)
Etage : Infralittoral
Nature du substrat : Fonds durs
Répartition bathymétrique : De la surface jusqu'à 35-40 m
Situation : Mer ouverte
Hydrodynamisme : Faible à très fort
Salinité : 37-38
Température : Oscillations saisonnières et journalières
Lumière : Forte luminosité
Régime trophique : Oligotrophe
Cet habitat est situé dans l'étage infralittoral qui s'étend depuis la zone où les émersions ne sont plus qu'accidentelles jusqu'à la limite au-delà de laquelle les magnoliophytes marines et les macrophytes photophiles ne peuvent plus survivre. Cette limite inférieure est conditionnée par la pénétration de la lumière, elle est donc extrêmement variable selon la topographie et la qualité de l'eau. Dans certaines zones d'eau très claire, elle peut descendre jusqu'à 35 à 40 m, alors qu'elle est limitée à seulement quelques mètres dans les zones les plus turbides.
Tous les substrats rocheux de l'étage infralittoral où règnent des conditions de lumière suffisantes sont recouverts par des peuplements extrêmement riches et variés de macrophytes photophiles en strates érigées et de macrophytes sciaphiles en sous-strate dont le développement est généralement favorisé par une bonne disponibilité en lumière.
L'habitat héberge une biocénose d'une grande richesse et d'une extrême complexité.
On distingue trois horizons :
- un horizon supérieur (de 0 à 1 m), où la lumière et l'énergie hydrodynamique sont fortes ;
- un horizon moyen (de 1 à 15 m), où les facteurs lumière et hydrodynamisme sont fortement atténués ;
- un horizon profond (de 15 à 40 m), où la lumière et l'hydrodynamisme sont extrêmement faibles.
À chacun de ces horizons correspondent des associations et des faciès bien caractéristiques, parmi ceux-ci les principaux sont :
Pour l'horizon supérieur :
- Association à Ericaria amentacea (Syn. Cystoseira amentacea var. stricta) / Ericaria mediterranea (Syn. Cystoseira mediterranea) (III.6.1.a.) en eau pure, mode agité, avec forte luminosité ;
- Association à Ericaria crinita (Syn. Cystoseira crinita), C. foeniculacea (Syn. C. discors) et C. compressa / C. crinitophylla (III.6.1.b.) en eau pure, mode calme, avec forte luminosité ;
- Association à Pterocladiella capillacea et Schottera nicaeensis / Botryocladia botryoides (III.6.1.q.) en eau pure, mode agité, avec lumière atténuée ;
- Association à Padina pavonica, Dictyotales, Halopteris scoparia (Syn. Stypocaulon scoparium) et Laurencia spp. / Anadyomene stellata (III.6.1.k.) en eau pure, mode calme, avec forte luminosité ;
- Association à Sargassum spp. (III.6.1.f.) en eau pure, mode agité, avec forte luminosité ;
- Association à Dictyopteris polypodioides (Syn. D. membranacea), Zonaria tournefortii (III.6.1.u.) en eau pure, mode agité, avec forte luminosité ;
- Association à Ellisolandia elongata (Syn. Corallina elongata) et Herposiphonia secunda (III.6.1.g.) en mode moyen, avec forte luminosité ;
- Faciès de surpâturage à algues calcaires encroûtantes et oursins (III.6.1.x.) en milieu perturbé ;
- Faciès à Mytilus galloprovincialis (III.6.1.w.) dans les zones à fort apport organique.
Pour l'horizon moyen :
- Faciès des grands Hydraires (III.6.1.z.).
Pour l'horizon inférieur :
- Association à Gongolaria montagnei (Syn. Cystoseira spinosa) et Gongolaria usneoides (Syn. Cystoseira usneoides) (III.6.1.e.).
Ce dernier horizon comporte des espèces provenant du Coralligène (IV.3.1.).
Sa dynamique saisonnière est forte. Cet habitat est certainement le plus riche et le plus diversifié de tous. Il est dominé par la végétation et sa dynamique est largement conditionnée par le cycle biologique des macroalgues. Suivant les associations, son maximum de développement se situe en hiver ou en été. Dans tous les cas, la plupart des macroalgues ayant un cycle annuel, les thalles tombent ou sont arrachés, entraînant une partie des épiphytes et de la faune.
Les associations et faciès correspondant à des conditions environnementales bien particulières, seules des modifications du tracé de la côte ou des altérations du milieu peuvent les faire évoluer.
L'habitat héberge de très nombreuses espèces.
Algues : Lithophyllum incrustans, Padina pavonica,Halopteris scoparia (Syn. Stypocaulon scoparium), Laurencia obtusa, Amphiroa rigida, Jania rubens, Ericaria amentacea (Syn. Cystoseira amentacea var. stricta), Codium bursa.
Cnidaires : Actinia equina, Anemonia sulcata, Eudendrium spp., Sertularella ellisii, Aglaophenia octodonta.
Mollusques : Acanthochitona fascicularis, Patella aspera, Vermetus triquetrus, Mytilus galloprovincialis.
Polychètes : Amphiglena mediterranea, Branchiomma lucullanum, Hermodice carunculata, Lepidonotus clava, Lysidice ninetta, Perinereis cultrifera, Platynereis dumerilii, Polyophthalmus pictus, Syllis spp.
Crustacés : Perforatus perforatus, Ampithoe ramondi, Dexamine spiniventris, Hyale spp., Acanthonyx lunulatus.
Echinodermes : Amphipholis squamata, Paracentrotus lividus. Paracentrotus lividus est une espèce réglementée
Amendement protocole Barcelone : Annexe III
Convention relative à la conservation de la vie sauvage et du milieu naturel de l'Europe (Convention de Berne) : Annexe III
Ces biocénoses se caractérisent par la présence de nombreuses algues photophiles recouvrant les substrats durs.
Sur les côtes rocheuses, le contact supérieur se fait avec la biocénose de la roche médiolittorale inférieure (RMI) (II.4.2.) dans laquelle remontent certaines espèces lorsque les conditions le permettent. Le contact inférieur se fait avec la biocénose coralligène (C) (IV.3.1.) avec parfois des échanges de populations au niveau de l'horizon inférieur.
La limite de l'horizon inférieur est parfois difficile à distinguer de la biocénose coralligène (C) (IV.3.1.).
Cet habitat est présent sur toutes les côtes rocheuses naturelles et sur les substrats solides artificiels des côtes de Méditerranée.
Structure : forêt algale/peuplements arborescents
Fonctions : nurseries, production primaire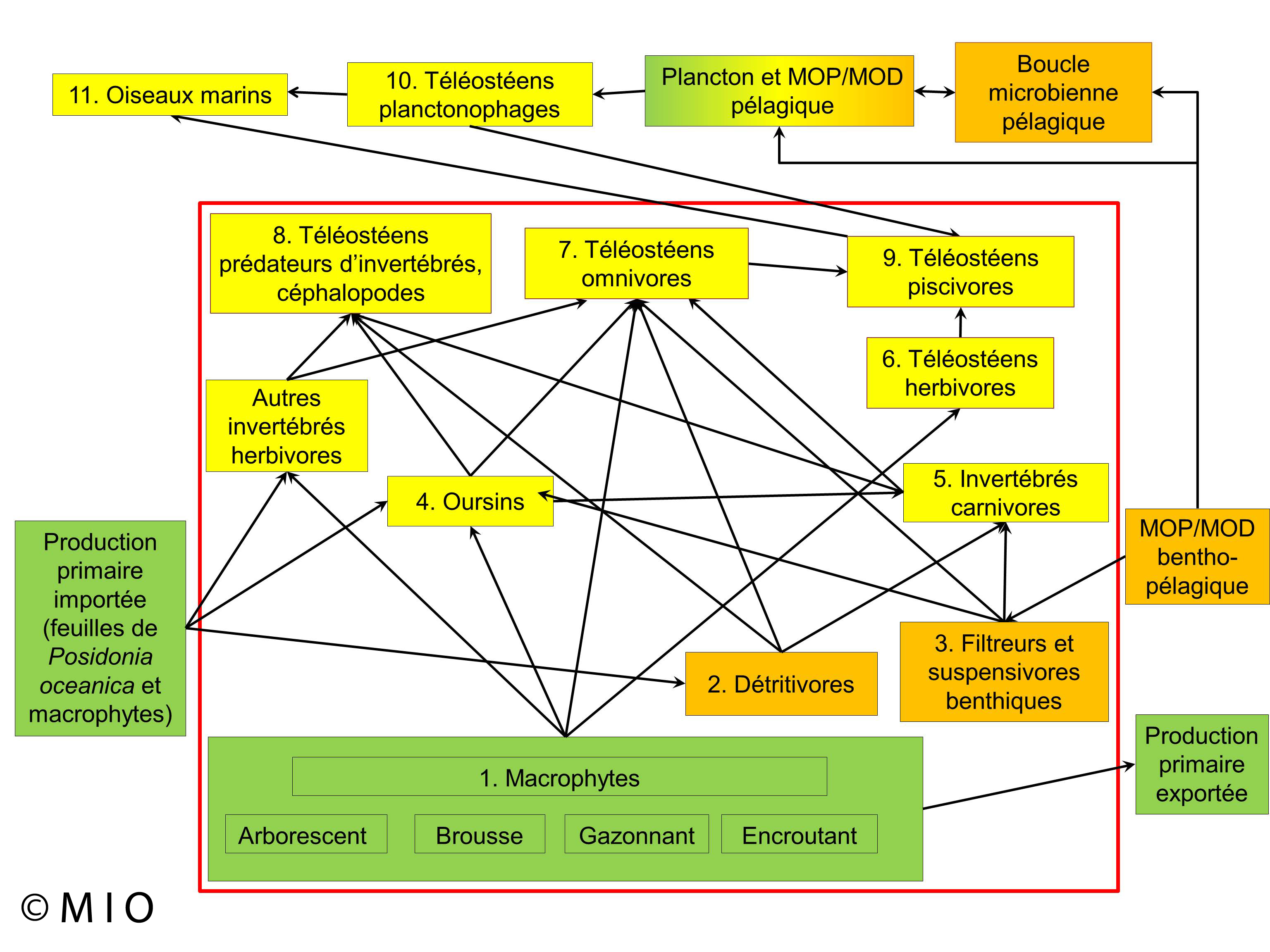
Figure 1 : Modèle conceptuel du fonctionnement de la biocénose des algues infralittorales traduit de Thibaut et al. (2017)
Cet habitat, extrêmement riche qualitativement et quantitativement, comprend plusieurs centaines d'espèces et sa production peut atteindre plusieurs kilogrammes par mètre carré. Le réseau trophique y est très complexe et ouvert sur les autres habitats par exportation d'organismes et de matériel organique. L'habitat intervient directement dans l'alimentation d'un grand nombre de poissons, soit de façon directe, soit de manière indirecte par la dispersion de détritus végétaux et animaux dans les autres fonds.
La production propre de cet habitat est relativement faible, en Méditerranée, elle se limite à l'exploitation des moulières naturelles. Les élevages de moules se font aussi, à ce niveau, sur des substrats artificiels.
Cette biocénose comprend des associations très sensibles à la qualité des eaux ; la Ericaria amentacea (Syn. Cystoseira amentacea var. stricta) est ainsi considérée comme un excellent indicateur de la qualité de l'eau et sa disparition est liée à l'accroissement de la pollution. Cette biocénose est aussi très sensible à la quantité de matières en suspension pour deux raisons fondamentales : les eaux turbides diminuent la photosynthèse et altèrent donc le peuplement algal, la sédimentation comble les microcavités entre les algues et élimine la petite faune cryptique. Elle est aussi fortement soumise à la pression des espèces introduites plus ou moins invasives (Caulerpa taxifolia, Stypopodium schimperi) qui peuvent l'altérer voire la détruire. L'ichtyofaune vivant au niveau de cette biocénose est diverse et riche ; elle est donc soumise à une forte pression de pêche professionnelle et de loisirs. Parmi les autres exploitations des composants de cette biocénose, on note la collecte des oursins et l'exploitation des moulières naturelles. Les élevages de moules se font aussi dans cet habitat sur des substrats artificiels.
Selon Thibaut et al. (2012), l’état de conservation de la roche infralittorale est bon pour l’horizon haut en région Provence-Alpes-Côte d'Azur et en Corse, dégradé sur la Côte des Albères. L’horizon inférieur est dégradé en région Provence-Alpes-Côte d'Azur et sur la Côte des Albères et bon en Corse. Les espèces les plus vulnérables de la roche infralittorale sont les espèces du genre Cystoseira qui sont sensibles au surpâturage, à la turbidité et à l’arrachage par les engins de pêche. Ces causes expliquent vraisemblablement la disparition de nombreuses espèces sur la Côte des Albères et le déclin de la plupart des espèces sur le continent. Des mesures urgentes de conservation sont nécessaires.
Michez N.
2016
La Rivière M., Michez N., Delavenne J., Andres S., Fréjefond C., Janson A-L., Abadie A., Amouroux J-M., Bellan G., Bellan-Santini D., Chevaldonné P., Cimiterra N., Derolez V., Fernez T., Fourt M., Frisoni G-F., Grillas P., Harmelin J-G., Jordana E., Klesczewski M., Labrune C., Mouronval J-B., Ouisse V., Palomba L., Pasqualini V., Pelaprat C., Pérez T., Pergent G., Pergent-Martini C., Sartoretto S., Thibaut T., Vacelet J., Verlaque M., 2021. Fiches descriptives des biocénoses benthiques de Méditerranée. UMS PatriNat éd., Paris : 660 pp. (Source)
Michez N., Dirberg G., Bellan-Santini D., Verlaque M., Bellan G., Pergent G., Pergent-Martini C., Labrune C., Francour P., Sartoretto S., 2011. Typologie des biocénoses benthiques de Méditerranée, Liste de référence française et correspondances. Rapport SPN 2011 - 13, MNHN, Paris, 50 p. (Source)
Michez N., Fourt M., Aish A., Bellan G., Bellan-Santini D., Chevaldonné P., Fabri M.-C., Goujard A., Harmelin J.-G., Labrune C., Pergent G., Sartoretto S., Vacelet J., Verlaque M. 2014. Typologie des biocénoses benthiques de Méditerranée Version 2.Rapport SPN 2014 - 33, MNHN, Paris, 26 pp. (Source)
Bensettiti F., Bioret F., Roland J. & Lacoste J.-P. (coord.) 2004. « Cahiers d’habitats » Natura 2000. Connaissance et gestion des habitats et des espèces d’intérêt communautaire. Tome 2 - Habitats côtiers. MEDD/MAAPAR/MNHN. Éd. La Documentation française, Paris, 399 p. + cédérom.
PNUE, PAM, CAR/ASP, 2007. Manuel d’interprétation des types d'habitats marins pour la sélection des sites à inclure dans les inventaires nationaux de sites naturels d’intérêt pour la Conservation. Pergent G., Bellan-Santini D., Bellan G., Bitar G. et Harmelin J.G. eds., CAR/ASP publ., Tunis, 199 pp.
THIBAUT T., BLANFUNE A., BOUDOURESQUE C.F., PERSONNIC S., RUITTON S., BALLESTEROS E., BELLAN-SANTINI D., BIANCHI C.N., BUSSOTTI S., CEBRIAN E., CHEMINEE A., CULIOLI J.M., DERRIEN S., GUIDETTI P., HARMELIN-VIVIEN M., HEREU B., MORRI C., POGGIALE J.C., VERLAQUE M., 2016. An ecosystem-based approach to assess the status of mediterranean shallow algae-dominated rocky reefs. Marine Pollution Bulletin. Elsevier, 117 (1-2) : 311 - 329.
Thibaut T., Blanfuné A., Markovic L. 2012. Biocénoses des fonds durs de l’infralittoral. Sous-région marine Méditerranée occidentale. Evaluation initiale DCSMM. MEDDE, AAMP, Ifremer, Ref. DCSMM/EI/EE/MO/22/2012, 11 pp.
 French Version
French Version