Typologie nationale des biocénoses benthiques de Méditerranée (NatHab-Med)
Etage : Infralittoral en Méditerranée occidentale
Nature du substrat : Rocheux
Répartition bathymétrique : Niveau moyen, soumis à des émersions et immersions
Situation : Mer ouverte
Hydrodynamisme : Faible à très fort
Salinité : Normale, pouvant supporter une légère dessalure
Température : Normale, variations en fonction de la température de l'air. La température de l’eau ne doit pas trop se refroidir en hiver.
Lumière : Constante
Régime trophique : Oligotrophe
Les cuvettes se caractérisent par une grande variation des conditions écologiques et sont parfois associées aux vermets, seul ce dernier cas de figure est traité ici. Les vermets se rencontrent dans l’étage infralittoral (Pérès, 1967 ; Pérès et Picard, 1964). En Méditerranée occidentale, ce faciès correspond à des enclaves infralittorales dans l’étage médiolittoral (PNUE‐PAM‐ CAR/ASP, 2007). Ces gastéropodes sessiles se développent à proximité du niveau moyen de mer, juste en dessous de la limite supérieure de l’étage infralittoral, là où le ressac est le plus actif. On peut aussi retrouver ces organismes plus en profondeur (5 - 6 m) sur des surfaces rocheuses dénudées par les oursins. La sédimentation de la zone est faible (Pérès, 1967 ; Pérès et Picard, 1964 ; PNUE‐PAM‐ CAR/ASP, 2007 ; Templado et al., 2015). En France métropolitaine, les cuvettes à vermets ne sont pas présentes. Elles ont été répertoriées par Molinier en 1955 au Cap Corse. Ces cuvettes ont existé mais les connaissances actuelles ne permettent pas d’affirmer qu’elles y soient toujours.
Dendropoma cristatum (autres noms qui ne sont plus valides : Vermetus cristatus, Dendropoma petraeum) construit des coquilles en tubes calcaires bruns formant de minces placages plus ou moins continus sur la roche ou s’agrége en bourrelets qui peuvent dépasser 10 cm d’épaisseur. Ces constructions organogènes, formées de coquilles de vermets morts et au-dessus de vermets vivants, peuvent atteindre plusieurs centaines de mètres d’envergure, comme en Israël (Safriel, 1974). Des espèces de l’étage infralittoral accompagnent les vermets (voir les espèces associées).
Ces formations organogènes constituent des substrats pour les espèces de l’horizon inférieur de l’étage médiolittoral. Il existe différentes formes :
- Sur les tombants rocheux subverticaux : des formes en corniche ou encorbellement (figure 1.A) en dessous du niveau moyen de la mer sont observées. Ce sont des bourrelets simples sur des roches dures, cristallines ou éruptives (Bellan-Santini et al., 1994). On les retrouve juste sous la corniche de Lithophyllum byssoides. En dessous de ces encorbellements, une structure organogène constituée d’algues calcaires sciaphiles peut être observée. Un exemple a été observé à Centurie, à l’ouest du Cap Corse (Molinier, 1960).
- Sur les roches en pente : des formations comblent l’angle formé entre la surface du plan d’eau et la paroi rocheuse (figure 1.B). Elles ont été observées sur de la granulite de Pasci-Pecora près d’Ajaccio et sur du gneiss à la calanque de Paragnani près de Bonifacio en Corse (Molinier, 1960).
- Sur une plate-forme faiblement inclinée terminée par un tombant submergé : il peut se former des concrétions à 2 endroits : au point de contact de la roche et de la surface et au-dessus du tombant. Au niveau du tombant, une sorte de margelle rétrécie se forme. Cette margelle est rétrécie au niveau de la base et s’élargie au sommet (figure 1. C). Ce phénomène a été observé au Cap Corse sur les schistes lustrés d’Albo et sur les grès de Javea sur les côtes espagnoles (Molinier, 1960).
- Sur les plates-formes subhorizontales : Forme en "trottoir" ou en "plate-forme" (figure 2). La structure type a été décrite en Sicile pour la première fois par Quatrefages en 1854. Cette forme correspond à une surface de corrosion horizontale développée dans la roche calcaire en place. Ces plates-formes ont également été observées sur les côtes d’Algérie près de Tipaza (Pérès et Picard, 1952). Sur la roche calcaire usée par l’érosion physico-chimique et biologique, les vermets peuvent coloniser les zones rendues disponibles. C’est pour cela que certaines formations peuvent être rencontrées plus en profondeur. Des flaques plus ou moins profondes de quelques centimètres peuvent être observées. Si les conditions de développement optimales des vermets ne sont pas réunies, les organismes se développent uniquement sur le rebord externe de ces formations. Les rebords externes de ces plates-formes ont tendance à s’élever à l’aide de piliers plus ou moins irréguliers. La bordure intérieure de la plate-forme, dépourvue de vermets protecteurs, se retrouve exposée à une dissolution naturelle et la zone se transforme en cuvette. Ce phénomène a été observé à Tipaza par Guilcher en 1954 et à Milazzo en Sicile. L’intérieur de ces cuvettes peut être colonisé par Vermetus triqueter, anciennement Vermetus gregarius et par Cladocora caespitosa (PNUE‐PAM‐ CAR/ASP, 2007). Les Cladocora caespitosa n’ont cependant pas été observées, en France, dans les cuvettes à vermets.
- En Méditerranée orientale, Israël et Crête, une autre forme en atoll a été observée. Ces atolls correspondent à des édifices arrondis et déprimés au centre. Ces formes sont comparables aux ‘boliers’ des Bermudes (Safriel, 1974 ; Kelletat, 1979).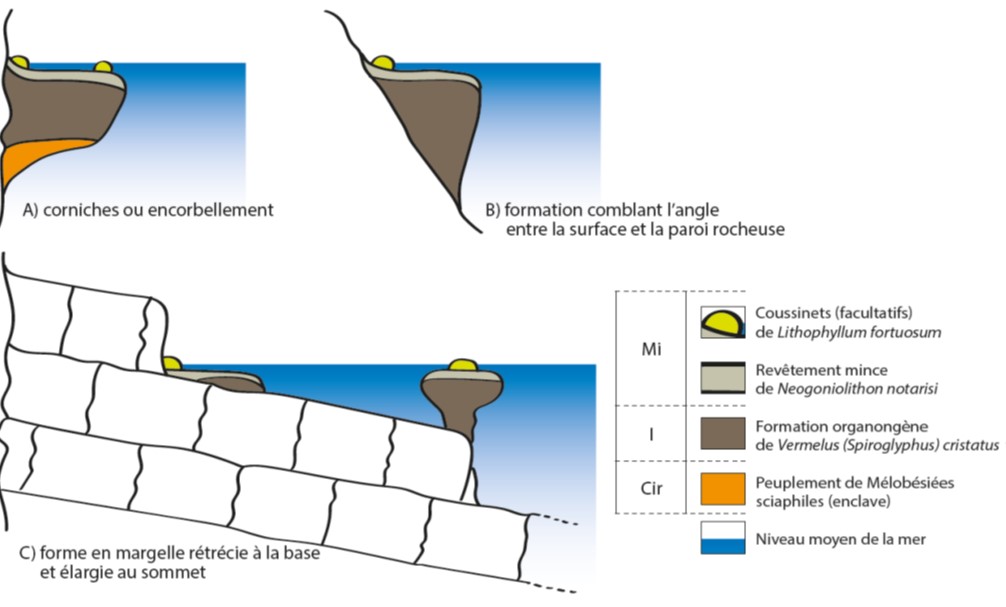
Figure 1 : Divers types de formations à Dendropoma cristatum (anciennement Vermetus cristatus). A) corniches ou encorbellement, B) formation comblant l’angle entre la surface et la paroi rocheuse et C) forme en margelle rétrécie à la base et élargie au sommet (Molinier, 1960)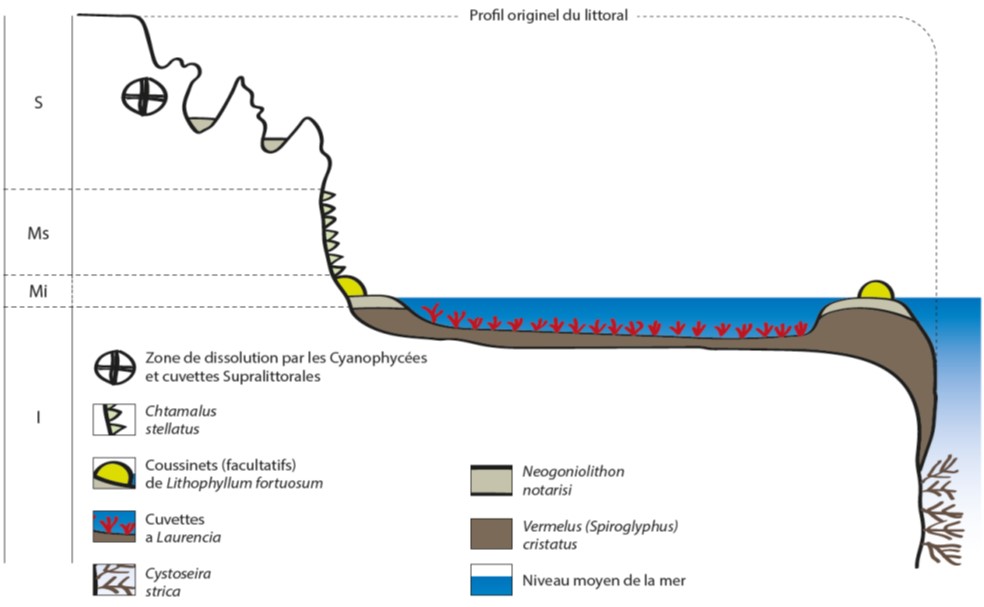
Figure 2 : Forme en trottoir ou en ‘plate-forme’ de Dendropoma cristatum (Pérès et Picard, 1952)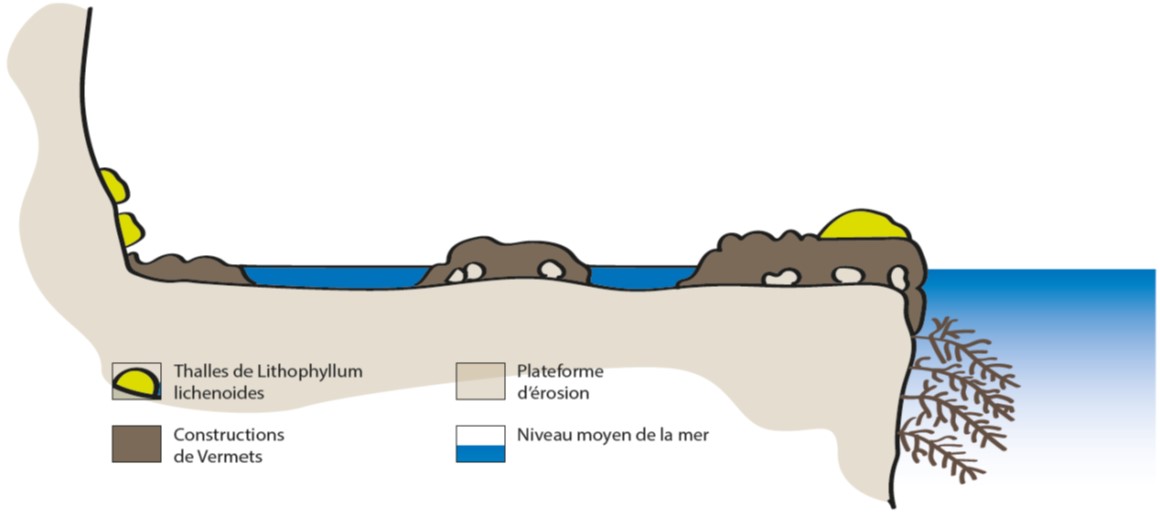
Figure 3 : Plate-forme à Vermets et ‘algues calcaires’ (Blanc et Molinier, 1955 dans Bellan-Santini et al., 1994)
Les Dendropoma cristatum sont fréquemment trouvés sur les rebords extérieurs des formations et les Vermetus triqueter sont plus rencontrés à l’intérieur des cuvettes.
Pas de dynamique particulière connue.
Dendropoma cristatum et Vermetus triqueter sont les espèces caractéristiques de l’enclave (PNUE‐PAM‐ CAR/ASP, 2007).
Des espèces de l’étage infralittoral accompagnent les vermets. Principalement, les Dendropoma cristatum sont associés à des algues calcaires comme Neogoniolithon brassica-florida et Lithophyllum byssoides. Il est fréquent de rencontrer, dans les tubes des vermets morts, le décapode Calcinus tubularis ; dans les cavités, l’oursin Arbaciella elegans (Algérie et Majorque) ; dans les ‘trous’, le foraminifère fixé Miniacina miniacea et dans les formations organogènes, le pélécypode Leiosolenus aristatus (Pérès et Picard, 1964 ; Bellan-Santini et al., 1994). Des mollusques tels que Lasaea adansoni et Patella aspera, l’éponge Sycon ciliatum et des annélides comme Perinereis macropus, ont été observés dans les encroûtements de vermets (Molinier, 1960). Des franges denses de Ericaria amentacea (Syn. Cystoseira amentacea var. stricta) ont été retrouvées au niveau du front des plates-formes. Diverses algues ont été observées sur les vermets à Farinole au Cap Corse comme Osmundea pinnatifida (Syn. Laurencia pinnatifida), Dictyota dichotoma et Gastroclonium clavatum (Molinier, 1960).
Les formes particulières des constructions biologiques des vermets sont caractéristiques. En général, Dendropoma cristatum se développe sur les rebords externes et les crêtes des platiers (PNUE‐PAM‐ CAR/ASP, 2007).
Association (encorbellement) à Lithophyllum byssoides (Syn. Lithophyllum lichenoides) (II.4.2.a.)
Association à Neogoniolithon brassica-florida et Lithophyllum byssoides (Syn. Lithophyllum lichenoides) (II.4.2.b.)
Les cuvettes assez profondes de la plate-forme, constituent des enclaves pour une flore et une faune appartenant à la biocénose des algues photophiles. Dans les endroits bien exposés, la limite avec l'horizon supérieur de l'étage infralittoral est parfois difficile à distinguer par le fait de la présence d'une série de petites mares en balconnets entre le niveau de la basse mer et celui de la plate-forme proprement dit (PNUE‐PAM‐ CAR/ASP, 2007).
Méditerranée occidentale : En France métropolitaine, cet habitat n’est jamais observé pour des raisons climatiques (refroidissement hivernal trop important pour le développement des vermets).
Au Cap Corse, à Ajaccio et dans les îles Lavezzi, une répartition irrégulière de ces organismes a été observée (Molinier, 1960). Au Cap Corse, Antonioli et al. (1999) ont considéré l’espèce éteinte de la zone et des observations directes devraient être réalisées pour confirmer cette disparition (Chemello, 2009 ; Chemello & Silenzi, 2011).
Des bio-constructions de vermets ont été observées dans d’autres zones de Méditerranée :
- Sardaigne, Italie, Sicile (Scuderi et al., 1998 ; Chemello 2009 ; Chemello et Silenzi, 2011)
- Malte (Azzopardi et Schembri, 1997)
- Tunisie et Algérie (Molinier et Picard, 1953)
- Crête, Grèce, Turquie, Syrie, Liban et Israël (Févert et Sanlaville, 1965 ; Safriel, 1974 ; Kelletat, 1979).
Une nouvelle espèce a été identifiée en 2016, Dendropoma lebeche. Elle est répartie sur les côtes de l'Atlantique et au sud-ouest de la Méditerranée (Minorque, les îles Baléares, le Delta de l’Ebre, le Cap Bon et les côtes rocheuses d’Algérie) (Templado et al., 2015).
En conclusion, pour ce qui est de la Méditerranée française, les vermets sont retrouvés uniquement en Corse, de façon très parcellaire et aucunes études récentes ne confirment leur présence actuelle.
Les vermets forment des bio-constructions en forme de plaquage ou de ‘trottoir’. Ces organismes créent un habitat par leur construction biogénique (appelé « espèce ingénieur »). En effet, il forme des structures ‘3D’ créant des habitats particuliers pour des organismes fixés ou mobiles.
L'intérêt de cet habitat consiste dans sa structure particulière comme un marqueur biologique des variations du niveau de la mer et comme un indicateur précis et fiable de la ligne de rivage. Il est fréquent de trouver en Méditerranée des trottoirs fossiles de l’holocène mais en Méditerranée française, seuls quelques trottoirs ont été retrouvés en Corse dans les années 1960. Il n’y a aucune étude récente affirmant leur présence. Ces trottoirs à vermets, à 0,80 m environ au-dessus de l'actuel niveau de la mer, ont été façonnés entre le début de l'époque hellénistique et le deuxième ou le troisième siècle de notre ère. Le trottoir à vermets constitue un élément majeur du paysage des côtes rocheuses, particulièrement en Méditerranée orientale. Les vermets sont également des espèces ingénieurs. Sous ces derniers, une ceinture dense ‘d’algues brunes’ du type Cystoseira peut-être observée. Ces macrophytes sont caractéristiques d’une ‘bonne qualité’ des eaux côtières (bien évidemment lorsque les conditions de développement de Cystoseira sont réunies). Le rebord externe du trottoir abrite une riche faune annélidienne et des organismes destructeurs, tels que les Sipunculiens (PNUE‐PAM‐ CAR/ASP, 2007). Etant un habitat accueillant de nombreuses espèces diverses, sa conservation est importante pour le maintien de la biodiversité.
Les plates-formes de vermets sont soumises à de nombreuses menaces anthropiques et leur lenteur d’édification les rendent encore plus vulnérables. Cet habitat est soumis à :
- La destruction par des aménagements sur la zone côtière : usines, complexes balnéaires et autres bétonnages, apports de sédiments et de débris.
- L'eutrophisation en zone urbaine qui entraine un envahissement des plates-formes par les Ulvales.
- L'action des pollutions d'origine diverse, comme celle provenant des mines d'amiante (par exemple l’usine de Canari en Corse), des usines de production des engrais phosphatés ou les hydrocarbures.
- L'utilisation des produits toxiques par les pêcheurs pour faire sortir les vers de leurs trous dans les flaques du trottoir et les utiliser comme appât.
- L'implantation volontaire ou accidentelle d'espèces introduites, dont le développement est rarement maîtrisé.
Les pollutions diverses agissant sur les bio-constructions de vermets, si elles ne sont pas maitrisées, peuvent engendrer des destructions quasi définitives de ces structures. Au Cap Corse, l’espèce a été considérée comme éteinte (Antonioli et al., 1999) et aucunes expertises récentes n’ont été réalisées afin de confirmer cette extinction.
Palomba L., Harmelin J.-G.
2016
La Rivière M., Michez N., Delavenne J., Andres S., Fréjefond C., Janson A-L., Abadie A., Amouroux J-M., Bellan G., Bellan-Santini D., Chevaldonné P., Cimiterra N., Derolez V., Fernez T., Fourt M., Frisoni G-F., Grillas P., Harmelin J-G., Jordana E., Klesczewski M., Labrune C., Mouronval J-B., Ouisse V., Palomba L., Pasqualini V., Pelaprat C., Pérez T., Pergent G., Pergent-Martini C., Sartoretto S., Thibaut T., Vacelet J., Verlaque M., 2021. Fiches descriptives des biocénoses benthiques de Méditerranée. UMS PatriNat éd., Paris : 660 pp. (Source)
Michez N., Fourt M., Aish A., Bellan G., Bellan-Santini D., Chevaldonné P., Fabri M.-C., Goujard A., Harmelin J.-G., Labrune C., Pergent G., Sartoretto S., Vacelet J., Verlaque M. 2014. Typologie des biocénoses benthiques de Méditerranée Version 2.Rapport SPN 2014 - 33, MNHN, Paris, 26 pp. (Source)
Antonioli, F., Chemello, R., Improta, S. et Riggio, S. (1999). Dendropoma intertidal reef formations and their paleoclimatological significance, NW Sicily. Marine Geology, 161, 155-170.
Azzopardi, L. et Schembri, P.J. (1997). Vermetid crusts from the Maltese Islands (Central Mediterranean). Marine Life, 7 (1-2), 7-16.
BELLAN-SANTINI D., LACAZE J.C., POIZAT C., 1994. Les biocénoses marines et littorales de Méditerranée, synthèse, menaces et perspectives. Collection Patrimoines Naturels. Secrétariat de la Faune et de la Flore/M.N.H.N., 19 : 1-246.
Bieler, R.; Gofas, S. (2012). Dendropoma cristatum (Biondi, 1859). In: MolluscaBase (2016). @ : http://www.marinespecies.org/aphia.php?p=taxdetails&id=195911, consulté le 25 avril 2016.
Boudouresque, C.F. (2015). Taxonomy and evolution of unicellular eukaryotes. Dans : Environmental microbiology : Fundamentals and applications. Microbial ecology. Bertrand, J.C., Caumette, P., Lebaron, P., Matheron, R., Normand, P. et Sime-Ngando, T. (éds.). Springer publisher : 191-257.
Chemello, R. (2009). Le bioconstruzioni marine in Mediterraneo. Lo stato delle conoscenze sui reef a vermeti. Biologia Marina Mediterranea, 16 (1), 2-18.
Chemello, R. et Silenzi, S. (2011). Vermetid reefs in the Mediterranean Sea as archives of sea-level and surface temperature changes. Chemistry and Ecology, 27 (2), 121-127.
Févert, M. et Sanlaville, P. (1965). Contribution à l'étude du littoral libanais. Méditerranée, 6 (2), 113-134.
Guilcher, A. (1954). Morphologie littorale du calcaire en Méditerranée occidentale (Catalogne et environs d'Alger). Bulletin de l'Association de géographes français, 31 (241-242), 50-58.
Kelletat, D. (1979). Geomorphologische Studien an den Küsten Kretas. Abhandlungen der Akademie der Wissenchaften in Gottingen. Mathematisch-Physikalische Klasse, 32, 1-105.
MOLINIER R., 1960. Étude des biocoenoses marines du Cap Corse. Vegetatio, 9 (3-5) : 121-312.
Molinier, R. (1955). Les plateformes et corniches récifales de Vermets. C. R. Ac. Sc., Paris 240 : 2 166. Dans Pérès, J. M. et Picard, J. 1964. Nouveau manuel de bionomie benthique de la mer Méditerranée. Station Marine d'Endoume, 31 (47), 137 pp.
Molinier, R. (1960). Etude des biocénoses marines du Cap Corse. Vegetatio, 9, 174-187.
Molinier, R. et Picard, J. (1953). Notes biologiques à propos d’un voyage d’étude sur les côtes de Sicile. Annales de l’Institut Océanographique, 28 (4), 163-188.
PERES J.M., PICARD J., 1964. Nouveau manuel de bionomie benthique de la Méditerranée. Rec Trav. Stat. Mar. Endoume, 31 (47) : 1-137.
PNUE, PAM, CAR/ASP, 2007. Manuel d’interprétation des types d'habitats marins pour la sélection des sites à inclure dans les inventaires nationaux de sites naturels d’intérêt pour la Conservation. Pergent G., Bellan-Santini D., Bellan G., Bitar G. et Harmelin J.G. eds., CAR/ASP publ., Tunis, 199 pp.
PÉRÈS J.M. 1967- The Mediterranean benthos. Oceanogr. Marine Biology Annual Review. 5. 449-553.
Pérès, J.M. et Picard, J. (1952). Les corniches calcaires d'origine biologique en Méditerranée occidentale. Recueil des Travaux de la Station Marine d’Endoume, 4, 2-33.
Quatrefages, A. (1854). Souvenir d’un Naturaliste. Tome 1, Charpentier, libraire-éditeur.
Safriel, U.N. (1974). Vermetid gastropods and intertidal reefs in Israel and Bermuda. Science, 186, 1113-1115.
Schiaparelli, S., Guidetti, P. et Cattaneo-Vietti, R. (2003). Can mineralogical features affect the distribution patterns of sessile gastropods? The Vermetidae case in the Mediterranean Sea. Journal of the Marine Biological Association of the UK, 83, 1267-1268.
Scuderi, D., Terlizzi, A. et Faimali, M. (1998). Osservazioni su alcuni tratti della biologia reproduttiva di vermeti biocostruttori e loro ruolo nella edificazione dei “trottoir”. Biologia marina mediterranea, 5 (1), 284-289.
Templado, J., Richter, A. et Calvo, M. (2015). Reef building Mediterranean vermetid gastropods: disentangling the Dendropoma petraeum species complex. Mediterranean Marine Science, 17 (1), 13-31.
 English Version
English Version