Typologie nationale des biocénoses benthiques de Méditerranée (NatHab-Med)
Etage : Circalittoral, souvent en enclave dans l’infralittoral
Nature du substrat : Rocheux ou bioconstruction
Répartition bathymétrique : 3 à 60 m
Situation : Mer ouverte
Hydrodynamisme : Le plus souvent réduit
Salinité : 38
Température : Variations saisonnières
Lumière : Faible luminosité
Régime trophique : Oligotrophe, apport faible mais régulier de nourriture
La biocénose des grottes semi-obscures (GSO) est principalement rencontrée sur les parties les plus extérieures des grottes (jusqu’à environ 40 m), au niveau des tunnels mais également au niveau des surplombs, fissures et parois verticales ombragées. Cette biocénose se trouve à la transition entre les fonds de substrats durs plus ou moins éclairés (selon la profondeur et l’orientation) où peuvent se développer des algues calcaires et les grottes obscures (GO) où l’environnement physique est très sélectif. Elle est présente sur la plupart des côtes rocheuses, particulièrement quand elles sont karstiques comme en Provence, de la Côte Bleue à Cassis.
Au niveau des grottes, cette biocénose est dépendante de la topographie de l’entrée de la grotte. Les différentes formes que peuvent avoir l’entrée des grottes vont influencer les facteurs abiotiques et donc la possibilité de colonisation par différentes espèces. Dans cet habitat, les facteurs tels que la lumière et l’hydrodynamisme sont réduits selon des gradients plus ou moins linéaires. Ceci entraîne une certaine stabilité du milieu et une représentation plus faible de certains groupes d’organismes (comme les filtreurs passifs par exemple). Une juxtaposition fréquente des faciès est également observée dans cette biocénose. La topographie du milieu et les évènements historique de recrutement en sont sans doute la cause (e.g. Bensettiti et al., 2004 ; PNUE, PAM, CAR/ASP, 2007 ; MNHN, 2012).
La variabilité de la biocénose est surtout d'ordre stationnel. Suivant la topographie du milieu et la modification des facteurs qui s'ensuit, on distingue un certain nombre d'aspects ou faciès :
- Faciès à Parazoanthus axinellae (IV.3.3.a.) : lorsque l'agitation des eaux est relativement élevée et l'éclairement encore significatif.
- Faciès à Corallium rubrum (IV.3.3.b.) : typique et fréquent par endroits, recouvre les parois des grottes, les cavités du concrétionnement coralligène et les surplombs semi-obscurs.
- Faciès à Leptopsammia pruvoti sous les surplombs et à l'entrée des grottes.
- Faciès à scléractiniaires Polycyathus muellerae, Caryophyllia inornata et Hoplangia durotrix : localisé dans les fissures ou les cavités des parois de grottes où l'obscurité est plus forte.
- Faciès à grands bryozoaires : près des entrées de grottes.
- Faciès d’appauvrissement liés à un hydrodynamisme plus intense avec abondance d’hydraires : Sertularella, Eudendrium.
- Faciès à Oscarella (jusqu’à 3 espèces pouvant la composer).
(e.g. Bensettiti et al ., 2004 ; PNUE, PAM, CAR/ASP, 2007 ; MNHN, 2012 ; Jean Vacelet, com. pers.)
La biocénose des grottes semi-obscures ne comprend que quelques rares algues sciaphiles, limitées à la zone la plus proche du milieu extérieur, et ne comprend donc en principe pas d’herbivores stricts. Le réseau trophique est donc constitué uniquement de filtreurs, de détritivores et de carnivores. Un degré de confinement se manifeste suivant un gradient qui va de l’extérieur vers l’intérieur de la grotte, avec une diminution des apports extérieurs, et un développement d’organismes fixés peu exigeants ou bien adaptés à l’exploitation d’une ressource trophique faible et aléatoire (e.g. Bensettiti et al., 2004 ; PNUE, PAM, CAR/ASP, 2007)
Les grottes semi-obscures sont peuplées par de nombreuses espèces d’invertébrés sessiles telles que des éponges, des scléractiniaires et des bryozoaires. Le Corallium rubrum et l’éponge Aplysina cavernicola sont deux espèces caractéristiques de cette biocénose (Laborel, 1961 ; Pérès et Picard, 1964).
Cette biocénose se situe généralement au niveau de l’entrée des grottes. Le substrat est composé de surplombs et de parois rocheuses abruptes au niveau desquels la lumière est fortement atténuée. Il est peuplé par de nombreuses espèces d'invertébrés sessiles caractéristiques, notamment à port dressé, assez colorées (voir espèces caractéristiques) (PNUE, PAM, CAR/ASP, 2007).
Suivant le gradient de lumière qui s'exprime en fonction de la profondeur et suivant l'éloignement de l'entrée des cavités, on trouve successivement la biocénose coralligène (C) (IV.3.1.), celle des grottes semi-obscures (GSO) (IV.3.3.) et celle des grottes et boyaux à obscurité totale (GO) (IV.3.4.) (Bensettiti et al. , 2004 ; PNUE, PAM, CAR/ASP, 2007). La biocénose des GSO peut également être en contact avec la biocénose des grottes médiolittorales (GM) (II.4.3.).
Lorsque cette biocénose occupe des cavités dans le concrétionnement coralligène (IV.3.1) ou la sous-strate des grands invertébrés arborescents (principalement des gorgones) du coralligène, elle peut ne pas être reconnue et être confondue avec cette dernière (Bensettiti et al., 2004 ; PNUE, PAM, CAR/ASP, 2007).
Cette biocénose est retrouvée au niveau de toutes les zones rocheuses, en particulier en enclaves ombragées en dehors des grottes. C’est d’autant plus le cas quand ces zones sont karstiques (calcaires) comme en Provence (Bouches-du-Rhône) de la côte Bleue à la Baie de Cassis (Bensettiti et al., 2004). En France, d’autres zones rocheuses propices sont rencontrées sur la côte des Albères, dans le Var (par exemple Parc National de Port Cros (Harmelin et al ., 2003)), la Côte d’Azur et Monaco, ainsi que dans l’ouest de la Corse (CREOCEAN, 2010). Elle peut exister occasionnellement au large de côtes sableuses quand des remontées rocheuses s’y prêtent (Languedoc, côte orientale de la Corse).
Les grottes semi-obscures sont des zones de transition entre le monde littoral éclairé et photophile et le monde cavernicole obscur. Ce sont également des zones servant de refuge pour toutes les proies potentielles de prédateurs visuels (y compris l’homme). Les variations nycthémérales de la faune mobile y sont donc importantes.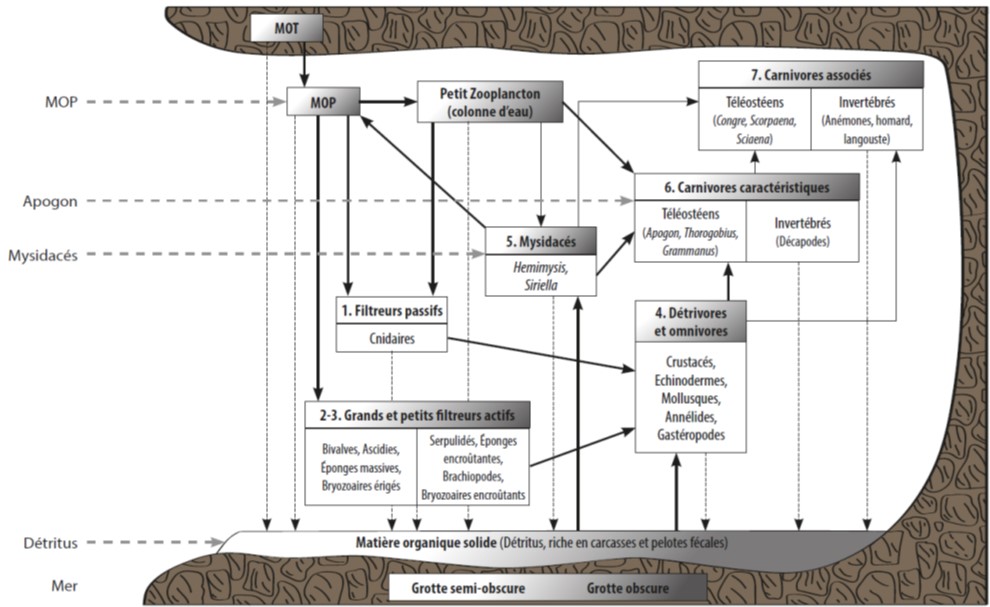
Figure 1 : Modèle théorique de la structure et du fonctionnement des écosystèmes des grottes sous-marine de Méditerranée. La différence entre les grottes semi-obscures (GSO) et les grottes obscures (GO) apparait en tête de chaque compartiment. Les flèches noires représentent l’intensité des flux de matière organique à travers les différents compartiments, avec les flèches noires pleines représentant les principaux flux. Les flèches en tirets noires verticales représentent la matière organique arrivant au fond de la grotte comme la matière organique d’origine terrestre (MOT), la matière en suspension particulaire (MOP), les pelotes fécales et les organismes morts. Les flèches en tirets grises horizontales représentent les principaux flux de matière entrant dans la grotte soit par advection des mouvements d’eau (pour la MOP et les détritus) soit par le biais des migrations circadiennes des Mysidacés et des Apogons (Rastorgueff et al., 2015)
Cette biocénose est extrêmement intéressante car elle comporte des espèces à haute valeur patrimoniale et/ou en danger ou menacées comme Corallium rubum, Aplysina cavernicola, Epinephelus marginatus, Homarus gammarus, Sciaena umbra, Scyllarides latus, Scyllarus arctus, Luria lurida, Schilderia achatidea et Zonaria pyrum. Cette biocénose représente à ce titre un modèle d’étude particulièrement intéressant de peuplements benthiques le long de gradients environnementaux et d'oligotrophie, à des profondeurs qui les rendent facilement accessibles aux observations. La conservation de cette biocénose passe par trois points : la surveillance de la qualité des eaux et de la pollution, en particulier de la charge particulaire, la gestion rigoureuse de la récolte du corail rouge, l’éducation des personnes pratiquant les activités sous-marines et le respect strict des réglementations (Bensettiti et al., 2004 ; PNUE, PAM, CAR/ASP, 2007).
Les entrées de grottes constituent des paysages de haute valeur esthétique. Elles sont donc fréquemment visitées par les plongeurs, notamment car elles sont riches en couleurs et faciles d'accès. Leur fréquentation exagérée peut, en provoquant une remise en suspension de la vase du plancher, une modification et une multiplication des contacts avec les organismes fixés, peut mettre en péril l'équilibre du peuplement.
Les anomalies de température liées aux changements globaux peuvent également affecter cette biocénose, en particulier le corail rouge et les éponges (Pérez et al., 2000 ; Laubier et al., 2003 ; Garrabou et al., 2009).
L'exploitation du corail rouge, de haute valeur marchande pour la bijouterie, est réglementée au niveau national et international, mais ces mesures de gestion doivent être strictement appliquées car le taux de croissance de cet octocoralliaire est très faible.
(PNUE, PAM, CAR/ASP, 2007 ; Bensettiti et al., 2004 ; MNHN, 2012).
Les faciès à Corallium rubum (IV.3.3.b), associé à cette biocénose, ont subi récemment des mortalités massives dont la cause le plus souvent évoquée est des anomalies thermiques. D'autres mortalités massives d’invertébrés ont été observées suite au réchauffement prolongé des étés 1999 et 2003 par exemple (Pérez et al., 2000 ; Garrabou et al., 2009). Si la méridionalisation de la Méditerranée continue, la biocénose des grottes semi-obscures risque de tendre vers un appauvrissement des espèces qui y sont présentes (e.g. Bensettiti et al., 2004 ; PNUE, PAM, CAR/ASP, 2007 ; MNHN, 2012)
Palomba L.,Chevaldonné P., Pérez T., Harmelin J.-G.
2016
La Rivière M., Michez N., Delavenne J., Andres S., Fréjefond C., Janson A-L., Abadie A., Amouroux J-M., Bellan G., Bellan-Santini D., Chevaldonné P., Cimiterra N., Derolez V., Fernez T., Fourt M., Frisoni G-F., Grillas P., Harmelin J-G., Jordana E., Klesczewski M., Labrune C., Mouronval J-B., Ouisse V., Palomba L., Pasqualini V., Pelaprat C., Pérez T., Pergent G., Pergent-Martini C., Sartoretto S., Thibaut T., Vacelet J., Verlaque M., 2021. Fiches descriptives des biocénoses benthiques de Méditerranée. UMS PatriNat éd., Paris : 660 pp. (Source)
Michez N., Dirberg G., Bellan-Santini D., Verlaque M., Bellan G., Pergent G., Pergent-Martini C., Labrune C., Francour P., Sartoretto S., 2011. Typologie des biocénoses benthiques de Méditerranée, Liste de référence française et correspondances. Rapport SPN 2011 - 13, MNHN, Paris, 50 p. (Source)
Michez, N., Fourt, M., Aish, A., Bellan, G., Bellan-Santini, D., Chevaldonné, P., Fabri, M.-C., Goujard, A., Harmelin, J.-G., Labrune, C., Pergent, G., Sartoretto, S., Vacelet, J. et Verlaque, M. 2014. Typologie des biocénoses benthiques de Méditerranée Version 2. Service du patrimoine naturel, Muséum national d'Histoire naturelle, Paris. SPN 2014 - 33: 26 pp. (Source)
BELLAN-SANTINI D., LACAZE J.C., POIZAT C., 1994. Les biocénoses marines et littorales de Méditerranée, synthèse, menaces et perspectives. Collection Patrimoines Naturels. Secrétariat de la Faune et de la Flore/M.N.H.N., 19 : 1-246.
Bensettiti F., Bioret F., Roland J. & Lacoste J.-P. (coord.) 2004. « Cahiers d’habitats » Natura 2000. Connaissance et gestion des habitats et des espèces d’intérêt communautaire. Tome 2 - Habitats côtiers. MEDD/MAAPAR/MNHN. Éd. La Documentation française, Paris, 399 p. + cédérom.
CREOCEAN, (2010). Inspection des grottes submergées et semi-submergées sur le littoral corse et propositions de mesures de gestion. DREAL de Corse, Dossier 1-10208E.
Garrabou J., Coma R., Bensoussan N., Bally M., Chevaldonné P., Cigliano M., Diaz D., Harmelin J.G., Gambi M.C., Kersting D.K., Ledoux J.B., Lejeusne C., Linares C., Marschal C., Perez T., Ribes T., Romano H., Serrano J., Torrents T., Zabala E., Zuberer S. et Cerrano A. (2009). Mass mortality in Northwestern Mediterranean rocky benthic communities: effects of the 2003 heat wave. Global Change Biology, 15 (5), 1090-1103.
HARMELIN J.G., VACELET J., VASSEUR P., 1985. Les grottes sous-marines obscures : un milieu extrême et un remarquable biotope refuge. Tethys, 11 (3-4) : 214-229.
Harmelin, J. G., Boury-Esnault, N., Fichez, R., Vacelet, J. et Zibrowius, H. (2003). Peuplement de la grotte sous-marine de l’Ile de Bagaud (Parc National de Port-Cros, France, Méditerranée). Scientific Reports of the Port-Cros Natural Park, 19, 117-134.
Laborel, J. (1961). Contribution à l'étude directe des peuplements benthiques sciaphiles sur substrat rocheux en Méditerranée. Recueil des Travaux de la Station Marine d’Endoume, 33 (20), 117-173.
Laubier, L., Pérez, T., Lejeusne, C., Garrabou, J., Chevaldonné, P., Vacelet, J., Boury-Esnault, N. et Harmelin, J.G. (2003). La Méditerranée se réchauffe-t-elle ? Marine life, 13 (1-2), 71-81.
MNHN. Service du Patrimoine Naturel (2012). Biocénoses des fonds durs du circalittoral. Sous-région marine Méditerranée occidentale. Evaluation initiale DCSMM. MEDDE, AAMP, Ifremer, Ref. DCSMM/EI/EE/MO/25/2012, 8 p.
PERES J.M., PICARD J., 1964. Nouveau manuel de bionomie benthique de la Méditerranée. Rec Trav. Stat. Mar. Endoume, 31 (47) : 1-137.
PEREZ T., GARRABOU J., SARTORETTO S., HARMELIN J.G., FRANCOUR P., VACELET J., 2000. Mortalité massive d’invertébrés marins : un événement sans précédent en Méditerranée nord-occidentale. C.R. Acad. Sci. Paris, Sciences de la Vie, 323 : 853-865.
PNUE, PAM, CAR/ASP, 2007. Manuel d’interprétation des types d'habitats marins pour la sélection des sites à inclure dans les inventaires nationaux de sites naturels d’intérêt pour la Conservation. Pergent G., Bellan-Santini D., Bellan G., Bitar G. et Harmelin J.G. eds., CAR/ASP publ., Tunis, 199 pp.
Rastorgueff, P.A., Bellan-Santini, D., Bianchi, C.N., Bussotti, S., Chevaldonné, P., Guidetti, P., Harmelin, J.G., Montefalcone, M., Morri, C., Pérez, T., Ruitton, S., Vacelet, J. et Personnic, S. (2015). An ecosystem-based approach to evaluate the ecological quality of Mediterranean undersea caves. Ecological Indicators, 54, 137-152.
 French Version
French Version