Typologie nationale des biocénoses benthiques de Méditerranée (NatHab-Med)
Etage : Circalittoral parfois en enclave à faible profondeur
Nature du substrat : Rocheux, concrétionnements, plancher souvent vaseux
Répartition bathymétrique : Pas de connaissance scientifique au-delà de 40 m de profondeur
Situation : Milieu confiné
Hydrodynamisme : Très faible sauf exception
Salinité : 38
Température : Saisonnalité tamponnée, dépendante de la topographie (grottes descendantes vs. ascendantes)
Lumière : Aucune
Régime trophique : Généralement très oligotrophe
La biocénose des grottes et boyaux à obscurité totale (GO) est retrouvée dans les cavités immergées dont le rapport entre la taille de l’entrée et la longueur totale est faible, ou dans lesquelles un obstacle important à la pénétration de la lumière est présent. On la retrouve surtout dans les réseaux karstiques ennoyés, mais parfois aussi dans des cavités de petite taille et des microcavités isolées dans les amas de pierres ou au sein de certains concrétionnements. Elles constituent des enclaves du domaine aphotique dans la zone littorale, en présentant des conditions environnementales très originales, proches de celles rencontrées sur la pente continentale. Les deux facteurs clés sont l'absence de lumière, qui exclut les organismes photosynthétiques, et le confinement, qui exclut les organismes à forte demande trophique. Le renouvellement de l’eau des chambres obscures est généralement très faible ou occasionnel et dépend de facteurs topographiques, bathymétriques et géographiques locaux. L’hydrologie et la composition biochimique sont généralement très stables et les conditions extrêmement oligotrophes, les dépôts sédimentaires sont extrêmement fins. La très forte diminution de l’apport trophique et de colonisateurs depuis l’extérieur entraîne une sélection drastique de la faune établie dans cet habitat. Le taux de recouvrement biologique des parois de cet habitat peut atteindre 50 à 80 % dans les zones les plus riches, mais peut être quasi nul dans les parties les plus confinées. Un enduit d’oxydes de fer et de manganèse peut-être observé sur les parois des grottes qui se retrouvent noircies. La présence d’espèces vivant normalement à plus grande profondeur (espèces bathyales) s’explique par le fait qu’elles trouvent dans cet habitat les conditions de lumière, de stabilité du milieu et de nourriture similaire à celles de la pente continentale et à la proximité de canyons profonds. Les variations de température y sont fortement tamponnées par rapport à l’extérieur, et dans les grottes à profil descendant (par exemple grotte des 3PP, Parc National des Calanques), le piégeage par densité d’eaux froides pendant la période estivale reproduit même les conditions stables et froides de température du bathyal méditerranéen (e.g. Vacelet et al., 1994 ; Harmelin et al., 1985 ; Harmelin, 1997 ; Bensettiti et al., 2004 ; PNUE, PAM, CAR/ASP, 2007 ; MNHN, 2012).
Les grottes sous-marines obscures, malgré des origines, tailles, configurations et expositions très diverses, présentent une grande unité de structure et de physionomie. Les différences spécifiques entre les GO peuvent venir du hasard du recrutement. La sélection des groupes trophiques et des groupes morphologiques ainsi que l'organisation spatiale sont régies par les conditions environnementales propres à chaque grotte et par la stochasticité des événements de colonisation. Ainsi une grande variabilité de peuplement est observée d’une grotte à l’autre (Bensettiti et al., 2004 ; PNUE, PAM, CAR/ASP, 2007 ; MNHN, 2012).
La biocénose des grottes obscures est particulièrement dépendante des apports énergétiques en provenance de l’extérieur et en propagules colonisatrices, ainsi que certains paramètres temporaires. La production y est très faible. Elle diminue avec le confinement et ne présente, en général, pas de cycle régulier. L'installation et la reconstitution du peuplement sont extrêmement lentes et aléatoires en raison de l'éloignement des sources exogènes de recrutement et de la rareté des apports énergétiques (Bensettiti et al ., 2004 ; PNUE, PAM, CAR/ASP, 2007 ; MNHN, 2012)
La biocénose des grottes et boyaux à obscurité totale est caractérisée par sa faune sessile dominée par des éponges majoritairement encroûtantes, des bryozoaires et des serpulidés, une diversité spécifique qui peut être élevée, mais une biomasse toujours faible. Les espèces caractéristiques de cette biocénose :
- Eponges : Discodermia polymorpha, Neophrissospongia nolitangere, Oopsacas minuta, Lycopodina hypogea, Merlia deficiens, Neoschrammeniella bowerbankii, Oscarella viridis, Oscarella microlobata, Plakina crypta, Plakina trilopha, Plakina jani, Pseudocorticium jarrei, Diplastrella bistellata, Hexadella cf. pruvoti, Myceliospongia araneosa, Thymosiopsis cuticulatus, Petrobiona massiliana
- Foraminifères : Discoramulina bollii ;
- Cnidaires : Ceratotrochus magnaghii ;
- Polychètes serpulides : Janita fimbriata, Filogranula annulata, Metavermilia multicristata, Vermiliopsis monodiscus ;
- Chaetognathes : Spadella ledoyeri ;
- Bryozoaires : Alcyonidium duplex, Glabrilaria pedunculata, Puellina corbula, Ellisina gautieri, Setosella cavernicola, Desmeplagioecia violacea, Annectocyma indistincta, Diaperoecia indistincta, Coronellina fagei, Setosella vulnerata ;
- Brachiopodes : Tethyrhynchia mediterranea, Argyrotheca cistellula ;
- Crustacés : Hemimysis speluncola, Hemimysis margalefi, Harmelinella mariannae, Stenopus spinosus, Herbstia condyliata ;
- Téléostéens : Grammonus ater, Gammogobius steinitzi, Thorogobius ephippiatus.
(Pérès et Picard, 1964 ; Bensettiti et al., 2004 ; MNHN, 2012)
Les fissures et les grottes ou parties de grottes, totalement dépourvues de lumière, où règnent des conditions de renouvellement de l’eau, de température et trophiques très particulières, rappelant les zones plus profondes, sont des critères de reconnaissance de cette biocénose. Le taux de recouvrement de la faune sessile peut y être très faible. La faune fixée adopte rarement un port dressé, les individus sont petits, ramassés, encroûtants, peu colorés. Les parois sont noircies par des enduits d’oxydes de fer et de manganèse assez caractéristiques de cette biocénose.
La biocénose des grottes obscures fait généralement suite à celle des grottes semi-obscures (GSO) (IV.3.3.) lorsqu'on s'enfonce dans une grotte une cavité sous-marine (Bensettiti et al., 2004 ). Cette biocénose peut également être en contact avec la biocénose des grottes médiolittorales (GM) (II.4.3.).
Dans les zones de transition, il est parfois difficile de déterminer les limites entre les 3 types de grottes : les grottes médiolittorales (II.4.3.), les grottes semi-obscures (IV.3.3.) et les grottes obscures (IV.3.4.) (Bensettiti et al., 2004 ).
Toutes les côtes rocheuses karstiques ou fracturées (côtes des Albères, côtes de Provence-Alpes-Côte d'Azur et côtes ouest de la Corse) sont susceptibles de présenter des éléments plus ou moins complets des grottes obscures avec une prédominance dans les zones karstiques (Bouches-du-Rhône) (Bensettiti et al., 2004).
Les grottes obscures, compte tenu des conditions particulières qui y règnent, sont des milieux refuges pour les organismes à faible compétitivité qui tolèrent les faibles ressources trophiques locales, contrairement à des organismes plus dynamiques. Cet effet refuge, par la stabilité du milieu, favorise la conservation d’éponges Lithistides. Les grottes constituent aussi des refuges pour des organismes cherchant à éviter les prédateurs visuels diurnes, comme par exemple des mysidacés cavernicoles, mais également de nombreux crustacés et téléostéens. Cette fonction refuge des grottes obscures peut être soit occasionnelle ou opportuniste, soit obligatoire (Harmelin et al., 1985 ; PNUE, PAM, CAR/ASP, 2007).
Dans cette biocénose des grottes obscures, la structuration de l’habitat se réalise à une très petite échelle. Dans certaines situations environnementales, des bryozoaires construisent des formations en nodules ou en crêtes parallèles sur les parois créant une certaine structuration. Ces espèces sont Onychocella marioni, Puellina radiata (Harmelin et al., 1985).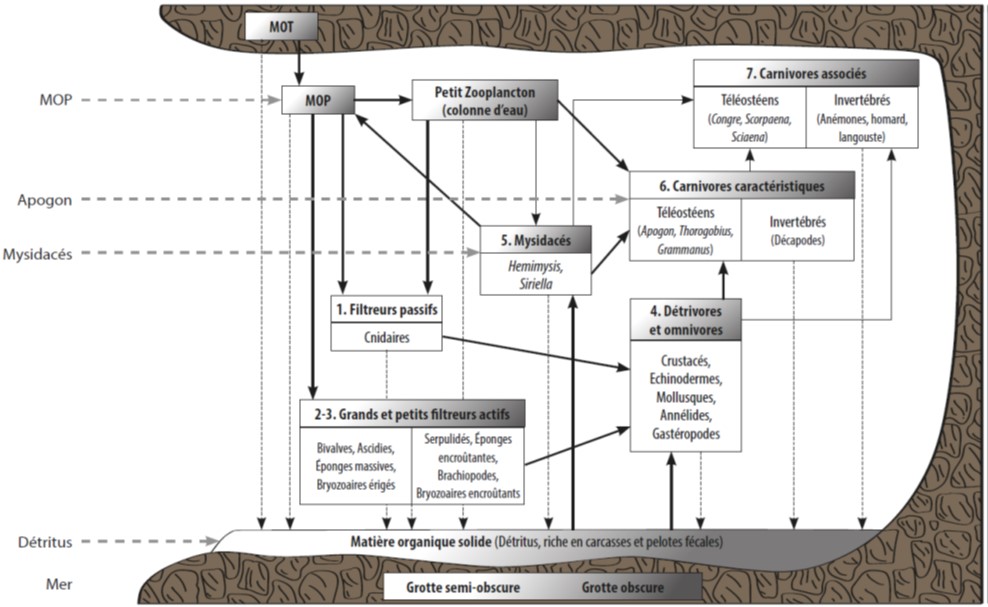
Figure 1 : Modèle théorique de la structure et du fonctionnement des écosystèmes des grottes sous-marine de Méditerranée. La différence entre les grottes semi-obscures (GSO) et les grottes obscures (GO) apparait en tête de chaque compartiment. Les flèches noires représentent l’intensité des flux de matière organique à travers les différents compartiments, avec les lèches noires pleines représentant les principaux flux. Les flèches en tirets noires verticales représentent la matière organique arrivant au fond de la grotte comme la matière organique d’origine terrestre (MOT), la matière en suspension particulaire (MOP), les pelotes fécales et les organismes morts. Les flèches en tirets grises horizontales représentent les principaux flux de matière entrant dans la grotte soit par advection des mouvements d’eau (pour la MOP et les détritus) soit par le biais des migrations circadiennes des Mysidacés et des Apogons (Rastorgueff et al., 2015)
Les grottes obscures sont des milieux refuges pour de nombreux organismes. Des espèces reliques y sont bien représentées (Petrobiona massiliana par exemple) et des éponges Lithistides ; d’autres y sont protégées de la prédation diurne (mysidacés cavernicoles comme Hemimysis margalefi). Des espèces bathyales peuvent vivre dans cette biocénose. En effet, les conditions trophiques, de lumière et de stabilité, parfois même de température, sont semblables à celles retrouvées dans l’habitat profond de ces organismes (par ex. Oopsacas minuta, Lycopodina hypogea). La plongée dans les grottes sous-marines donne ainsi accès à des études biologiques et écologiques uniques. La plongée de loisir, quant à elle, s’aventure rarement au-delà de la biocénose des GSO.
Le réchauffement global des eaux peut affecter les peuplements de ces milieux confinés. Chevaldonné et Lejeusne (2003) ont notamment montré le remplacement d’une espèce de mysidacés Hemimysis speluncola par H. margalefi, plus thermotolérante, dans les grottes de la région de Marseille.
La résilience de ces peuplements face aux perturbations naturelles et anthropiques est très faible. Leur trop forte fréquentation par les plongeurs pourrait mettre en péril l’équilibre de la biocénose en modifiant la circulation de l’eau, l’entrée de particules et par l’accumulation de bulles et la multiplication des contacts avec les organismes, mais ces dangers concernent essentiellement la biocénose des GSO, beaucoup plus fréquentée (Bensettiti et al., 2004).
La bonne gestion de cet habitat passe par deux séries de mesures : la surveillance de la qualité des eaux et de la pollution, en particulier de la charge en matières organiques et la gestion de la fréquentation et l’éducation des personnes pratiquant des activités sous-marine (Bensettiti et al., 2004 ; PNUE, PAM, CAR/ASP, 2007).
L’étude des grottes a vraiment commencé il y a une cinquantaine d’années, avec l’utilisation de la plongée en recherche scientifique. L’étude de la faune, de sa biologie et des facteurs environnementaux restent à approfondir. La biocénose des grottes obscures est a priori sensible à des changements locaux (e.g. pollution, espèces introduites) ou globaux (e.g. réchauffement) qui restent à évaluer (Bensettiti et al., 2004 ; MNHN, 2012, Harmelin J.-G., com. pers.).
Palomba L., Chevaldonné P., Harmelin J.-G.
2016
La Rivière M., Michez N., Delavenne J., Andres S., Fréjefond C., Janson A-L., Abadie A., Amouroux J-M., Bellan G., Bellan-Santini D., Chevaldonné P., Cimiterra N., Derolez V., Fernez T., Fourt M., Frisoni G-F., Grillas P., Harmelin J-G., Jordana E., Klesczewski M., Labrune C., Mouronval J-B., Ouisse V., Palomba L., Pasqualini V., Pelaprat C., Pérez T., Pergent G., Pergent-Martini C., Sartoretto S., Thibaut T., Vacelet J., Verlaque M., 2021. Fiches descriptives des biocénoses benthiques de Méditerranée. UMS PatriNat éd., Paris : 660 pp. (Source)
Michez N., Dirberg G., Bellan-Santini D., Verlaque M., Bellan G., Pergent G., Pergent-Martini C., Labrune C., Francour P., Sartoretto S., 2011. Typologie des biocénoses benthiques de Méditerranée, Liste de référence française et correspondances. Rapport SPN 2011 - 13, MNHN, Paris, 50 p. (Source)
Michez, N., Fourt, M., Aish, A., Bellan, G., Bellan-Santini, D., Chevaldonné, P., Fabri, M.-C., Goujard, A., Harmelin, J.-G., Labrune, C., Pergent, G., Sartoretto, S., Vacelet, J. et Verlaque, M. 2014. Typologie des biocénoses benthiques de Méditerranée Version 2. Service du patrimoine naturel, Muséum national d'Histoire naturelle, Paris. SPN 2014 - 33: 26 pp. (Source)
Bensettiti F., Bioret F., Roland J. & Lacoste J.-P. (coord.) 2004. « Cahiers d’habitats » Natura 2000. Connaissance et gestion des habitats et des espèces d’intérêt communautaire. Tome 2 - Habitats côtiers. MEDD/MAAPAR/MNHN. Éd. La Documentation française, Paris, 399 p. + cédérom.
Bonhomme P., Ganteaume A., Bellan G., Cadiou G., Emery E., Clabaut P., Bernard G., Hervé G., Bourcier M., Boudouresque C.F., 2005. Etude et cartographie des biocénoses marines des calanques de Marseille à Cassis, y compris l’archipel de Riou. Phase 3 : Rapport final. Contrat GIS Posidonie-IFREMER-COM/GIP des Calanques. 1-156.
Chevaldonné, P. et Lejeusne, C. (2003). Regional warming-induced species shift in north-west Mediterranean marine caves. Ecology Letters, 6, 371-379.
FICHEZ R., 1990. Decrease in allochthonous organic inputs in dark submarine caves, connection with lowering in benthic community richness. Hydrobiologia, 207 : 61-69.
HARMELIN J.G., 1997. Diversity of bryozoans in a Mediterranean sublittoral cave with bathyal-like conditions: role of dispersal processes and local factors. Mar Ecol Prog Ser, 153: 139-152.
HARMELIN J.G., VACELET J., VASSEUR P., 1985. Les grottes sous-marines obscures : un milieu extrême et un remarquable biotope refuge. Tethys, 11 (3-4) : 214-229.
Harmelin, J.G. (1985). Bryozoan dominated assemblages in Mediterranean cryptic environments. Bryozoa: Ordovician to Recent, 135-143.
LABOREL J., VACELET J., 1958 – Etude des peuplements d’une grotte sous-marine du golfe de Marseille. Bulletin de l’Institut Océanographique de Monaco, 1120 :1-20
Lejeusne, C. et Chevaldonné, P. (2005). Population structure and life history of Hemimysis margalefi (Crustacea: Mysidacea), a ‘‘thermophilic’’ cave-dwelling species benefiting from the warming of the NW Mediterranean. Marine Ecology Progress Series, 287, 189-199.
MNHN. Service du Patrimoine Naturel (2012). Biocénoses des fonds durs du circalittoral. Sous-région marine Méditerranée occidentale. Evaluation initiale DCSMM. MEDDE, AAMP, Ifremer, Ref. DCSMM/EI/EE/MO/25/2012, 8 p.
PERES J.M., PICARD J., 1964. Nouveau manuel de bionomie benthique de la Méditerranée. Rec Trav. Stat. Mar. Endoume, 31 (47) : 1-137.
PNUE, PAM, CAR/ASP, 2007. Manuel d’interprétation des types d'habitats marins pour la sélection des sites à inclure dans les inventaires nationaux de sites naturels d’intérêt pour la Conservation. Pergent G., Bellan-Santini D., Bellan G., Bitar G. et Harmelin J.G. eds., CAR/ASP publ., Tunis, 199 pp.
Rastorgueff, P.A., Bellan-Santini, D., Bianchi, C.N., Bussotti, S., Chevaldonné, P., Guidetti, P., Harmelin, J.G., Montefalcone, M., Morri, C., Pérez, T., Ruitton, S., Vacelet, J. et Personnic, S. (2015). An ecosystem-based approach to evaluate the ecological quality of Mediterranean undersea caves. Ecological Indicators, 54, 137-152.
Vacelet, J. et Boury-Esnault, N. (1996). A new species of carnivorous sponge (Demospongiae: Cladorhizidae) from a Mediterranean cave. Dans : Willenz P. (Ed.), Recent Advances in Sponge Biodiversity Inventory and Documentation. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Biologie, 66 (Suppl.), 109-115.
Vacelet, J., Boury-Esnault, N. et Harmelin, J.G. (1994). Hexactinellid cave, a unique deep-sea habitat in the scuba zone. Deep Sea Research Part I: Oceanographic Research Papers, 41 (7), 965-973.
 French Version
French Version